单倍群C-M8(HaplogroupC-M8)又称为单倍群C1a1(HaplogroupC1a1),HaplogroupM...

单倍群C-M8(Haplogroup C-M8)又称为单倍群C1a1(Haplogroup C1a1),Haplogroup M105, Haplogroup M131,属于C-M130的下游支系。
| 中文名 | 外文名 | 分 布 | 先祖血统 |
|---|---|---|---|
| 单倍群C-M8 | Haplogroup C-M8 | 中国,朝鲜半岛,日本 | M130 → CTS11043 |
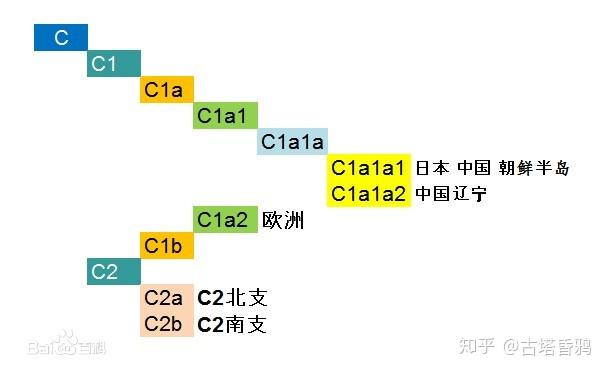
整个C单倍群分为 C1(C-F3393)和C2(C-M217),C1则分为C1a和C1b两支,C1a则又分为两支C1a1(C-M8)和C1a2。C1a在世界范围内是极其罕见的,主要是分布在东亚(C1a1)或欧洲(C1a2)。常说的C单倍群,在中国主要指的是C2,C2和C1,C1a的关系的,简单的归纳如下图:
起源
C-M8单倍群根据yfull计算,大概诞生于4万6千年前,是个古老的单倍群,有多古老,4万6千年前,东亚的主流单倍群N,O,R,Q还没有从各自的上游分出来。C1a1可以说是瓶颈期最长的父系单倍群,长达4万年之久(formed 46600 ybp - tmrca 4600 ybp ),C-M8下面积累了差不多470个SNP,瓶颈期相当严重,C1a1应该是较早进入东亚的单倍群,和他的亲戚C1a2一样,扩散较早,被后来的单倍群人群覆盖,在欧亚大陆的两端呈碎片化分布。
分布
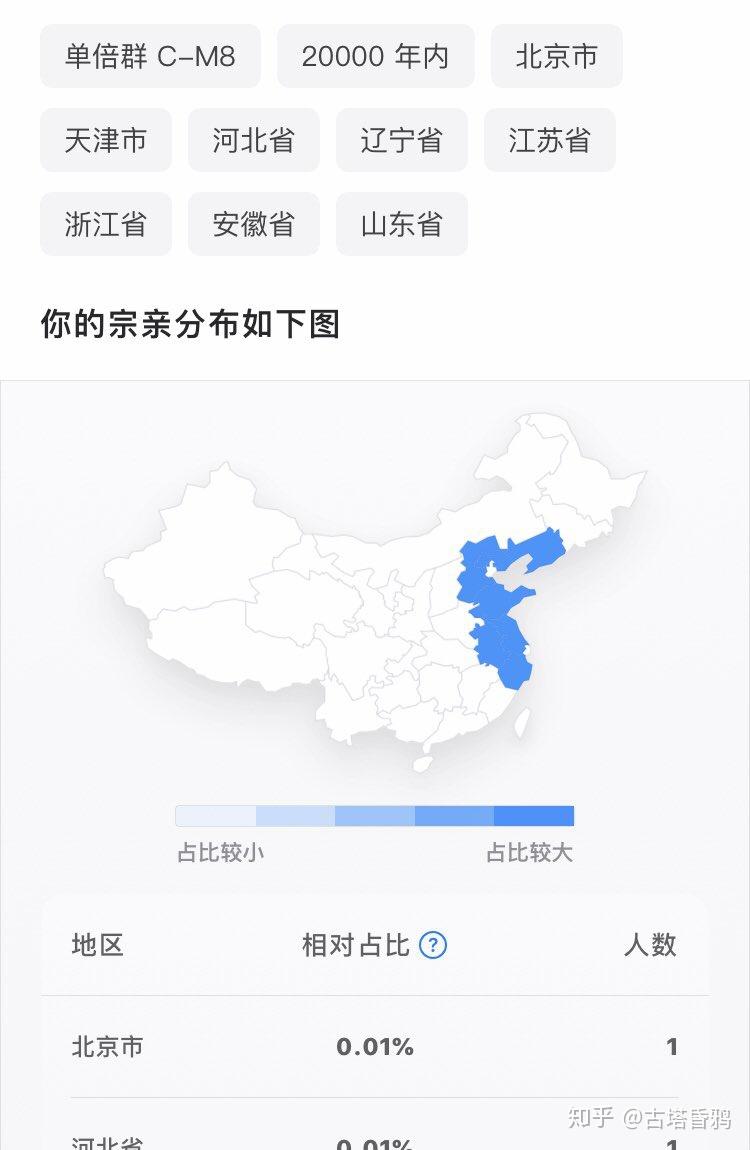
C1a1只分布在东亚的中国,朝鲜,韩国,日本,可以说是最稀少的东亚本土单倍群,中国北方沿海各省的比例在0.01%左右。在中国辽宁汉族,山东汉族,河北汉族,北京汉族,江苏汉族,浙江汉族,安徽回族,吉林延边朝鲜族(STR数据),徐州汉族(STR数据)检测到了C1a1单倍群。
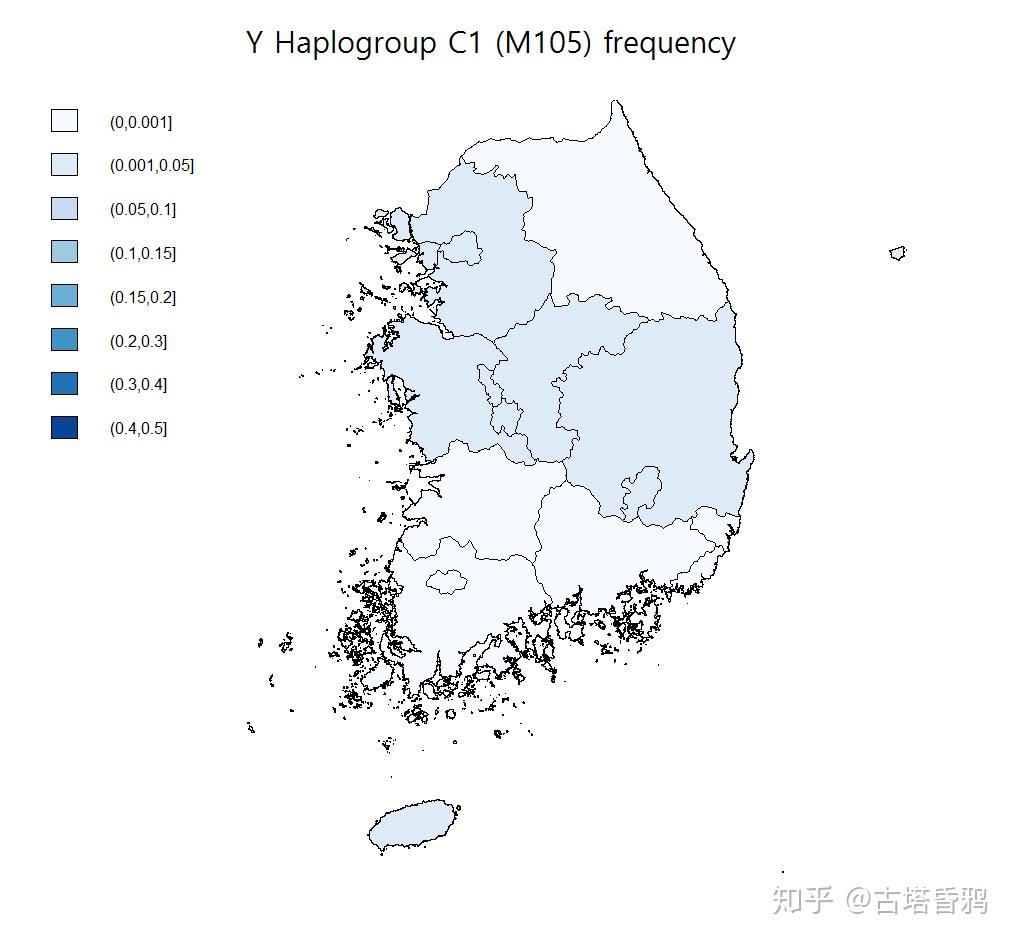
北朝鲜的一份123人的样本中发现2例C1a1样本,韩国的各个不同人群检测中也经常发现C1a1类型,整个韩国估计平均在0.5%左右,不到1%。1722 例韩国KRGDB样本中 C1a1-M8 样本 8例,早期支系 3例。
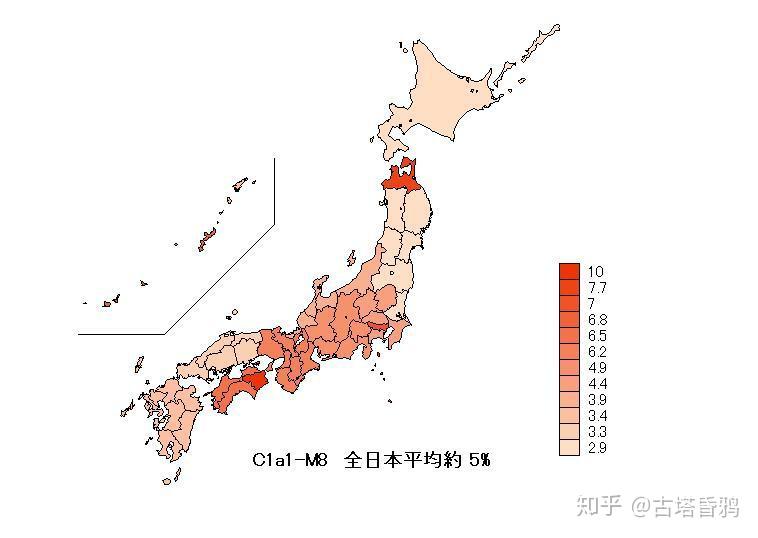
日本平均5.7% (采样于北海道,本州,四国,九州,冲绳的1299例STR数据中74例C1a1),最多的地方是四国地区,德岛县最高达到10%,东京7%,冲绳6.8%,大概的趋势就是中间多,两边少,越往日本东部北部越少,西部九州,东部太平洋沿岸,北海道最低。
日本国内的分布频率
德岛 10%
冲绳 6.8% [4.4%-8%]
本州 4.9% (青森 7.7%, 东京 7.0%, 静冈 4.9%)
九州 0%-3.8%
北海道 2.9%
阿伊努人 0%
结合其他单倍群的数据来看,D1b高的地区,C1a1都不高,而O1b高的地方,C1a1都不低,特别值得注意的是,N单倍群高的地方C1a1都高,N单倍群全日本平均0.8%到1%,而N单倍群最高的地区是 青森 7.7%,其次是的德岛地区5.7%。
当初在青海德令哈蒙古族中也发现了3个C1a1-M8的样本,复旦大学的文章,用的是Taqman分型,而且那套探针里明确有M8,M105的位点。
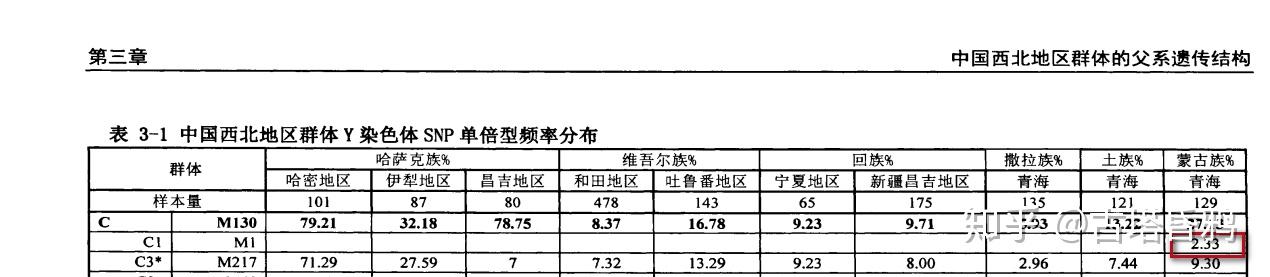
根据familytreedna的信息,在蒙古国,哈萨克斯坦,乌兹别克斯坦也发现了M105,也就是C1a1-M8,但是没看到具体的数据。
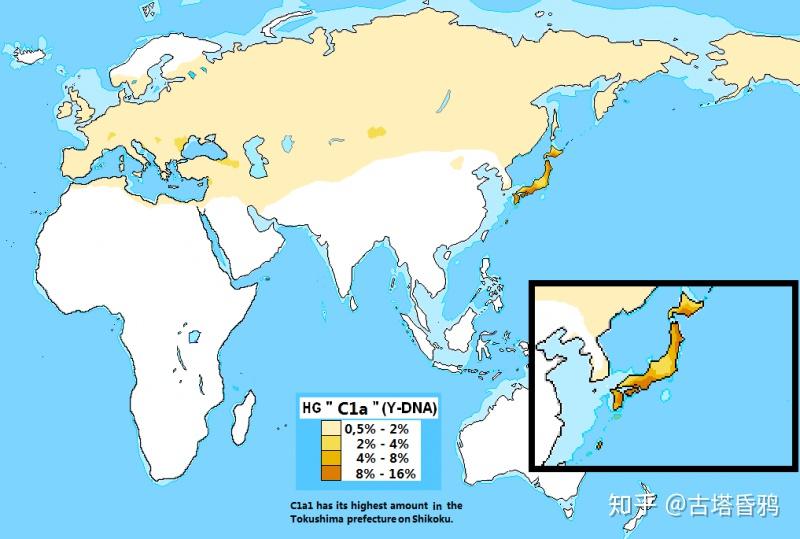
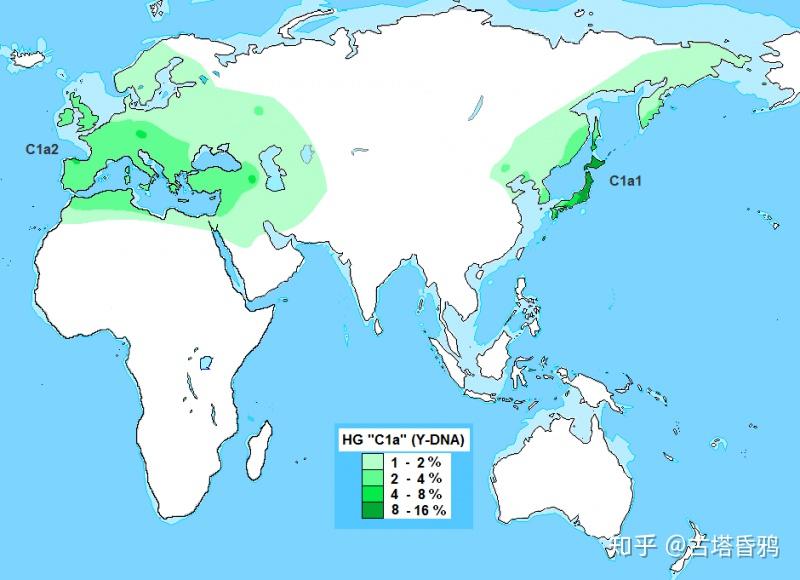
以下两张C1a1分布概念图来源于wiki
谱系树

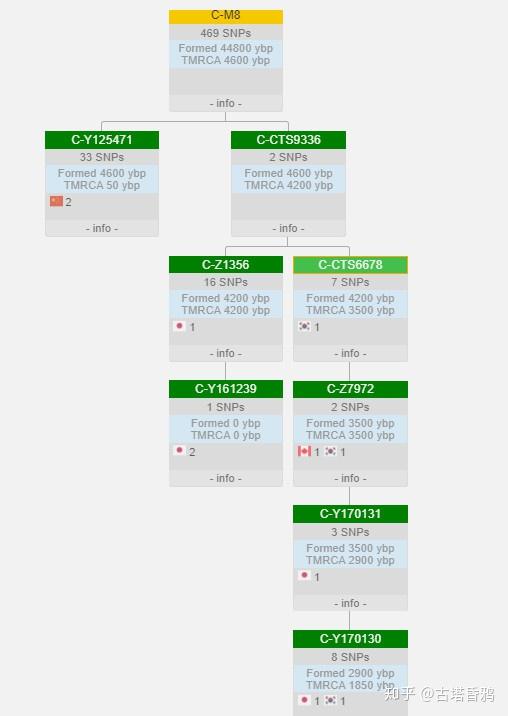
迁移路线
由于分布地域狭窄(中国,朝鲜半岛,日本列岛,冲绳群岛),分布频率较低,关于C1a1这个支系进入东亚的路线还是个谜,C1a1族群作为旧石器时代欧亚大陆北部的采集狩猎人群,活动范围较大,结合和C1a1最近的支系C1a2在欧洲,和现代人的分布频率,跟随猛犸象,披毛犀等大型猎物,沿着河流,湖泊迁徙,走北线进入东亚的可能性较大。日本崎谷満老师认为,C1a1人群经过伊朗,通过阿尔泰山,再通过的朝鲜半岛进入日本列岛。
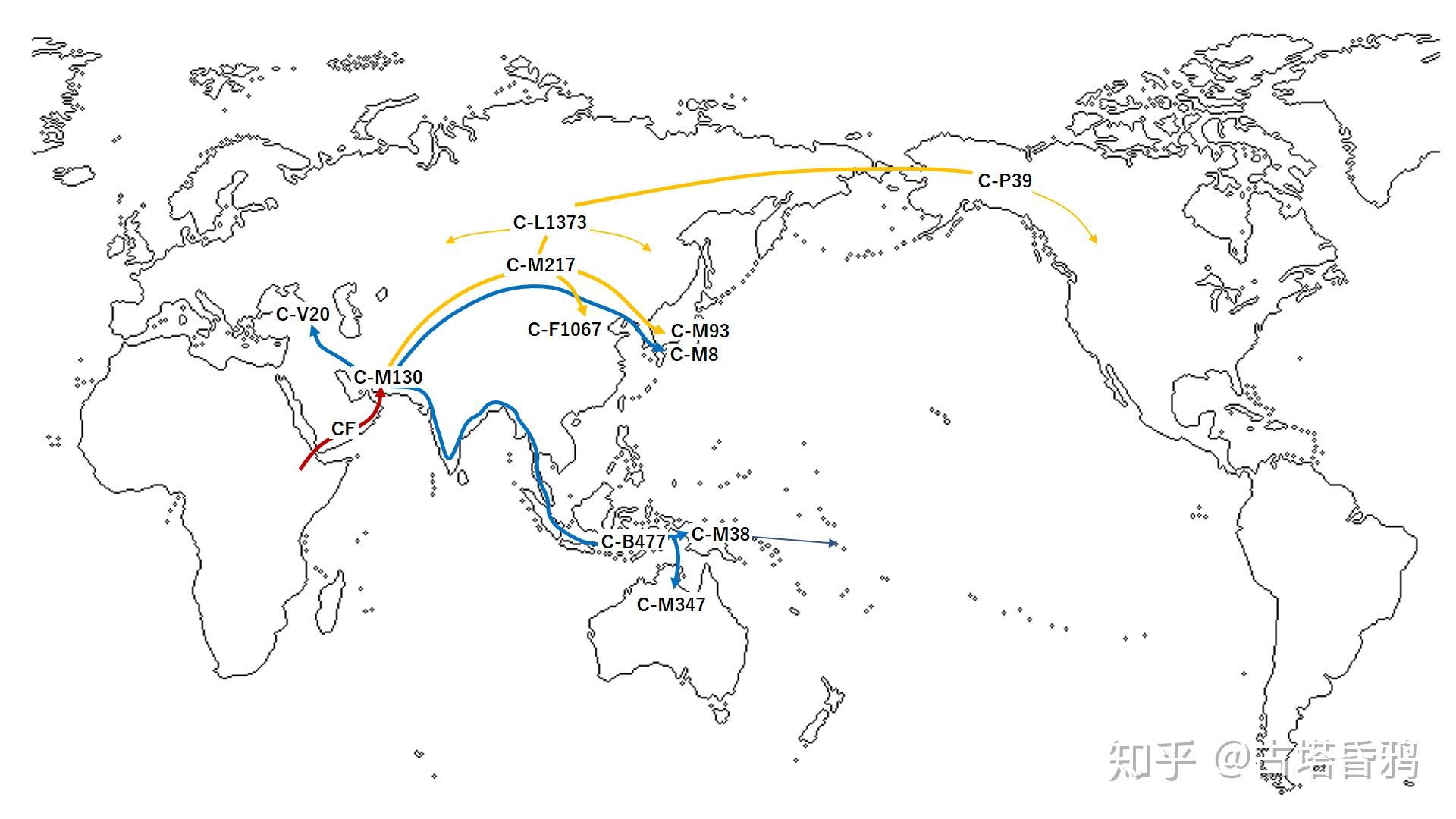
最近英文的Wiki 增加了下面的部分,引用的是 剑桥大学的文章,推测 北海道下面的青森的 大平山元I遗址(dai Yamamoto I Site)和 C1a1有关。认为进入日本的C1a1是通古斯民族。


古样本情况
最近一篇论文,在欧洲伊比利亚半岛13000年前的猎人古样本中发现了C1a1。期待以后进一步验证,因为这个位置很有可能是C1a2。

东亚的父系C1a1-M8族群最有可能就是山顶洞人的后裔。山顶洞遗址最新测年是距今3.5万年到3.8万年,这个期间正好是父系C1,C1a,C1a2族群活动的旺盛期,结合山顶洞遗址出来的装饰品和欧洲旧石器C1,C1a2古人遗址的相似性,再结合山顶洞古人和欧洲旧石器古人头骨基本一致的特点,认为山顶洞古人族群的父系是C1a2的东亚亲戚C1a1-M8的可能性很大。三万,四万年前,东亚地区体现奥瑞纳文化,格拉维特文化,马格德林文化的遗址,很有可能是C1a1人群留下的。
北京人和山顶洞人有延续性吗,和我华夏族人有连续性吗?www.zhihu.com弥生时代日本鸟取县青谷上寺地遗址,有4例样本父系有检测结果,其中两例为C1a1-M8。
常见QA
Q:阿伊努人的Y染色体类型是C1a1-M8 A:在现在的日本全国各地的阿伊努人中没有发现任何一例C1a1,阿伊努人中最大比例的单倍群是D1b,超过80%以上。
Q:冲绳的港川古人是C1a1(C1-M8)A:港川人并没有检测,也就没有所谓的检测的结果了,和C1a1(C1-M8)也没有任何关系。
Q:C1a1是日本固有的,特有的单倍群
A:从现在的结果看,朝鲜半岛平均0.5%,中国也有零星发现。C1a1这个单倍群是日本固有的单倍群这种说法是不准确的,特别是上游特有支系只发现在韩国和中国,日本数量虽然多,但类型集中,而都是下游类型。如果C1a1是日本固有特有的单倍群成立的话,不如说整个C1a(C1a1,C1a2)都是日本特有的类型好了,毕竟这个世界上绝大部分C1a都在日本。
讨论
C1a1分布地域狭窄,而且比例很低,高通样本更少,基本对该单倍群的研究几乎没有,基本都是靠想象,一般争议较大的就是迁移路线和进入日本的时间。之前的C单倍群按照C1,C2,C3分类的时候,认为日本的C1是从台湾,经过冲绳,进入日本,就是所谓的南路线,而且中国,朝鲜半岛的零星样本,是从日本扩散出去的的。C1a1的扩散时间和绳文时代的开始的时间一致(12000年以前),认为C1a1是日本绳文人。这些都是有正式论文支持的。


但是仔细一看就会发现问题,所谓的12000年,就是根据10STR的几个日本数据推算出来的,而且也是根据STR画的树形认为日本的C1和印度的C*更近,而且C1更多的都是在南半球,南面的类型更丰富,所以判断C1是从南亚走南线过来的。最近几年分子人类学的发展,确认了东亚的C1a1和欧洲的C1a2更近。
南半球的C1虽然多,但是都是C1b的下游。而且中国和朝鲜半岛的SNP的确认,发现日本的C1a1虽然数量多,但是集中在几个下游支系,暂时没有发现上游支系,中国和朝鲜半岛虽然数量少,但是都出现了上游支系,而且上游下游都有,类型比较丰富。
冲绳的C1a1比例较大,很多人认为冲绳是C1a1的源头,但是实际上以德岛为主的四国地区比例最多,人数最多,而且类型丰富。
分析一篇论文中所有的冲绳的STR,发现冲绳样本和Yfull上的样本的STR都很接近,都是日本列岛的常见类型,初步判断是几千年,从日本列岛南下的人群。冲绳是C1a1族群扩散的最南端。
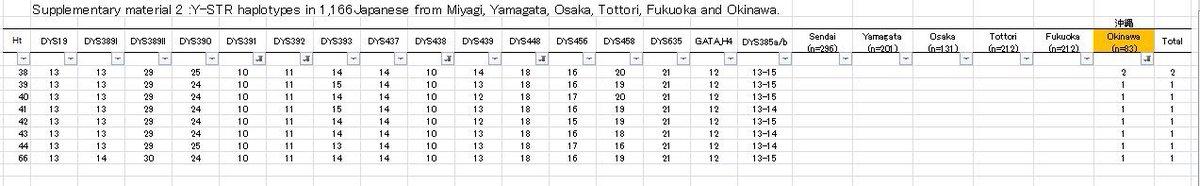
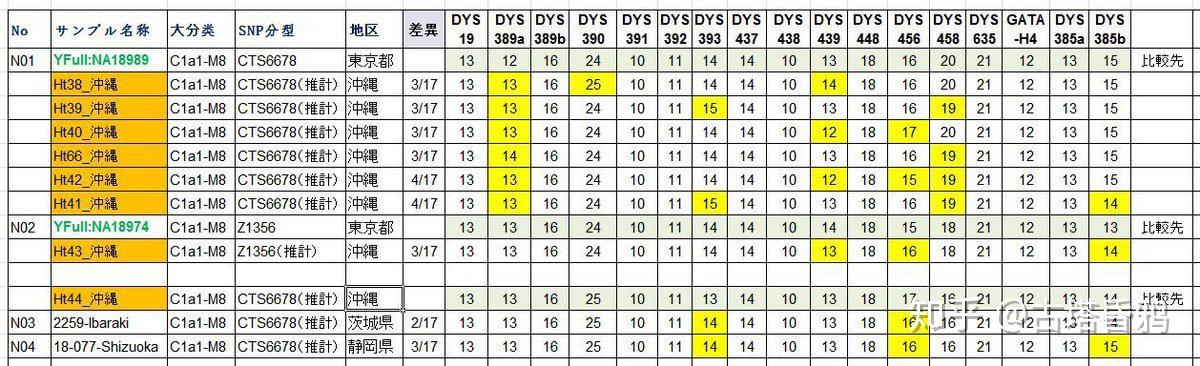
日本琉球大学正式否定了冲绳人南来的可能性,现代冲绳人都是从日本列岛扩散而来。

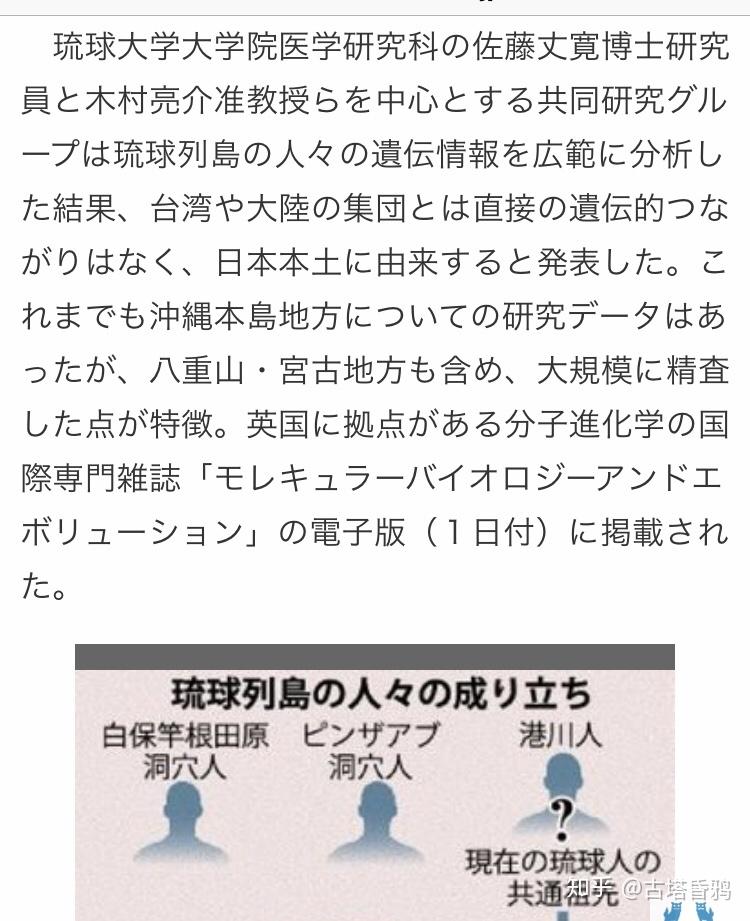
C1a1族群是否是绳文人,严格意义上不是,但是也可以说,因为绳文时代太长,弥生时代太短。就算3000年内进入日本列岛(弥生时代(公元前300-公元250年))也可以认为是绳文人。一般来想日本的绳文人的父系是D1b应该没有问题,但是C1a1瓶颈期太长,树形和D1b相差太大,而且现代人C1a1比例高的地区都是弥生成分多的地方,而且阿伊努人中至今没有发现C1a1,无论怎么看都是C1a1人群和D1b混合较晚,而且混合的十分不均匀。
这里要说一下,绳文人不等于早死,从出土的古人样本上分析,日本列岛的绳文人的寿命不短。根据最近从9处不同地点出土的 65具绳文时代人骨的最新检测结果得出,65岁以上的个体占到 32.5%,以前的研究结果 [縄文人=早死] 的结论是不对的,古代绳文人的寿命还是很长的,形質人類学的专家,考古学者也认同这个最新的结论,进行这项研究的是 圣玛丽安娜医科大学 讲师 长冈朋人(人类学·古人口学),作为文部科学省的科学研究费的成果,最近在《月刊考古学杂志》的临时增刊号上发表了这一结果。

所以说如果C1a1族群较早进入日本,不会有长达四万年的瓶颈期,也会和D1b混和的较均匀。
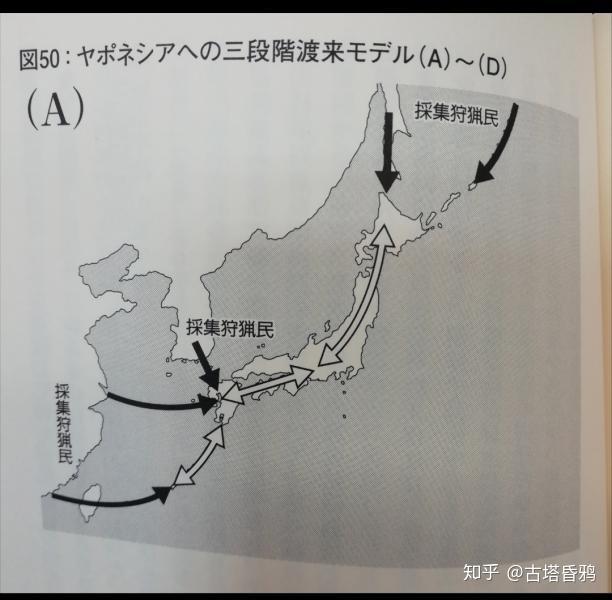
根据 齐藤成也 老师的三阶段理论,认为C1a1人群是第二批,第三批进入日本列岛的可能性很大。
第一阶段(狩猎采集人群)(约4万年前到4400年前的旧石器时代到绳文时代的中期)
第一批渡来人群从欧亚大陆各地通过各种路线流入日本。特别是,在1万2000年前日本一直是大陆相连,因此,容易进行进入。这批渡来人群主要以狩猎采集为主,在常染色体与现在大部分的东亚人有着很大不同。各个路线进入日本的各集团发展成的日本人被认为就是我们通常所说的绳文人。
第二阶段(捕鱼,园耕民)(约4400年前-3000年前) 绳文时代后期,第二批渡来人群流入。虽然这与绳文人和dna有很大的差异,但与后述的第三批渡来民也有所不同。斋藤认为,这个集团是居住在中国黄海沿岸的“海之民”,并以捕鱼为主要的狩猎采集者或狩猎采集和农耕为一体的园耕民。第二批渡来集团在日本列岛中心地区和第一批的绳文人混血,但在北海道、南西群岛、东北地区没有造成影响。
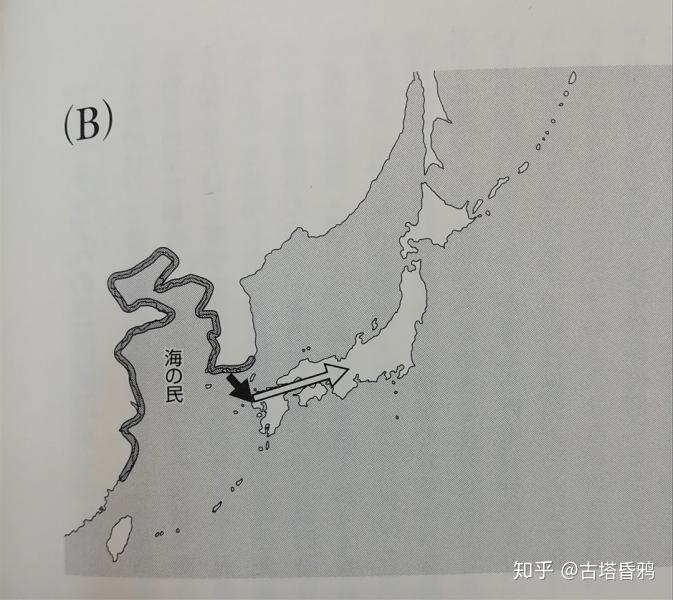
第三阶段 前期(农耕民) (约3000年前-1700年前) 通过朝鲜半岛的稻谷为主的渡来人群流入,引进了水田稻作。该集团主要是在北九州、畿内和连接关东的日本中心轴,在其他地方并没有产生较大的影响。特别是北海道和南西群岛,对东北的影响几乎没有的。
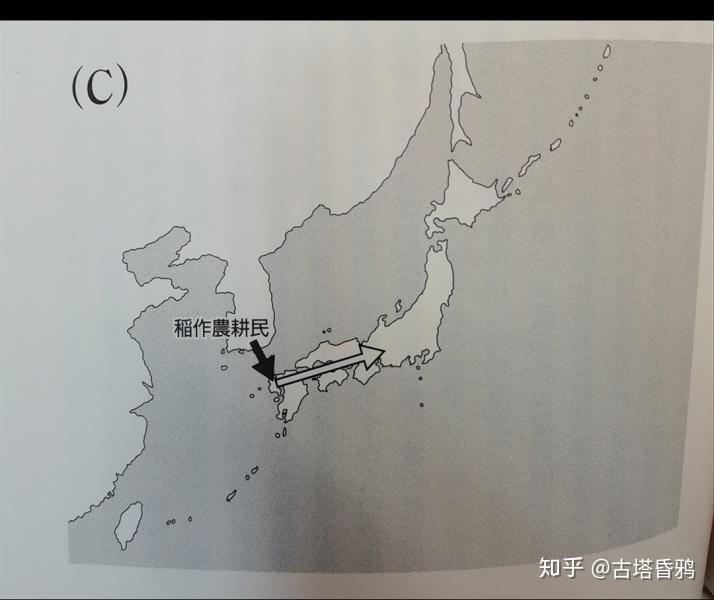
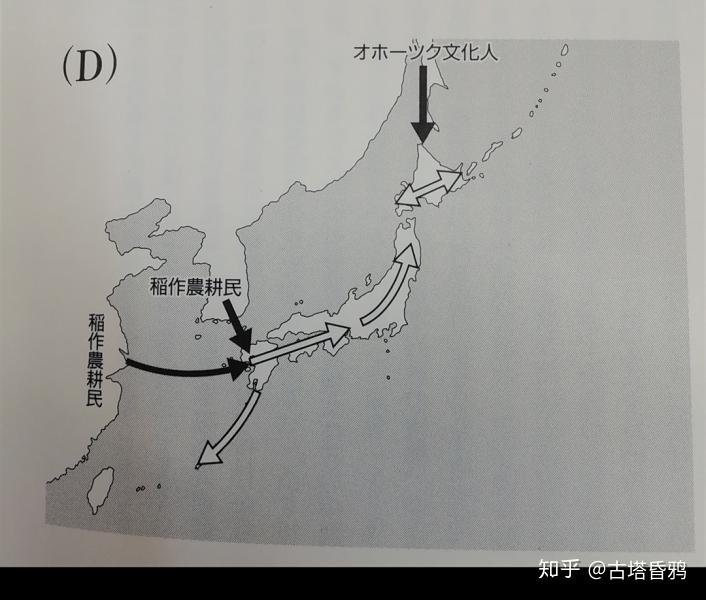
第三阶段 后期 (约1700年前-现在) 政治的中心转移到日本中部(京都,大阪附近),不仅是朝鲜半岛,现在的中国上海周边也有一些外来人群流入。进入古坟时代后,居住在日本东北地区的第一阶段(旧石器绳文人)的大部分人移居到北海道,因此,以第二阶段(捕鱼人群或园耕民)的子孙为中心的人们住到了日本的东北部。在南西群岛,古斯克时期(12世纪~15世纪),以从南九州到第二阶段的后裔为中心的集体移民、混血,在江户时代,与第三阶段集团的混血儿也进行了。在北海道,从古坟时代到平安时代,鄂霍次克文化人群(通古斯女真人群)开始和绳文人开始交流混血,江户时代以后,这些人又与日本本土的日本人也进行了混血。
关于C1a1现存支系的扩散路线无外乎下面几种。
1,台湾ー>冲绳ー>日本本岛ー>朝鲜半岛ー>中国。
2,北海道ー>日本本岛ー>朝鲜半岛ー>中国
3,中国ー>朝鲜半岛ー>日本本岛ー>冲绳
4,朝鲜半岛分别向中国,日本扩散。
我个人的看法,3,4的可能性更大,2的可能性最低,1期待更多日本检测结果的公布再继续进一步分析。
结尾语
现在关于C1a1的各种推论,证据链还很弱,本文中尽量把和C1a1有关的信息整理一下,也许有些是不完善,不正确的,主要是样本太少,期待未来更多的中日韩高通样本的出现来逐步揭开C1a1的种种谜团。









如果认为本文对您有所帮助请赞助本站